
Introduction
Staphylococcus aureus remains one of the most clinically challenging bacterial pathogens, responsible for conditions ranging from superficial skin infections to life-threatening bacteremia, endocarditis, and osteomyelitis. The mortality rate for complicated S. aureus bacteremia can reach 65–70%, and up to half of staphylococcal pneumonia isolates are now classified as methicillin-resistant S. aureus (MRSA), underscoring the urgent need for therapeutic approaches beyond conventional antibiotics.
Antibody discovery targeting S. aureus surface antigens has emerged as a promising strategy to address antibiotic resistance. By leveraging the specificity of monoclonal antibodies (mAbs) against defined virulence factors, researchers can neutralize immune evasion mechanisms, block bacterial adhesion to host tissues, disrupt nutrient acquisition, and directly counter antibiotic resistance determinants. This article examines the scientific rationale and preclinical progress for five key S. aureus antigen targets: Staphylococcal protein A (SpA), iron-regulated surface determinant B (IsdB), clumping factor A (ClfA), S. aureus surface protein A (SasA), and penicillin-binding protein 2a (PBP-2a).
The S. aureus Antigenic Landscape
- aureus expresses an extensive repertoire of cell wall-anchored (CWA) proteins and secreted virulence factors that collectively enable colonization, immune evasion, tissue destruction, and nutrient scavenging. Among these, the microbial surface components recognizing adhesive matrix molecules (MSCRAMMs)—including ClfA, SasA, and IsdB—mediate attachment to host extracellular matrix proteins, while SpA functions primarily as an immune evasion factor by binding immunoglobulin Fc and Fab regions. PBP-2a, encoded by the acquired mecA gene, represents a resistance determinant rather than a classical virulence factor, yet it is arguably the most clinically consequential surface antigen in MRSA isolates.
The functional diversity of these antigens presents both a challenge and an opportunity for antibody-based intervention. Targeting a single antigen risks leaving compensatory pathways intact; conversely, the availability of multiple validated targets opens the door to combination strategies that may achieve more robust protection than monotherapy approaches.
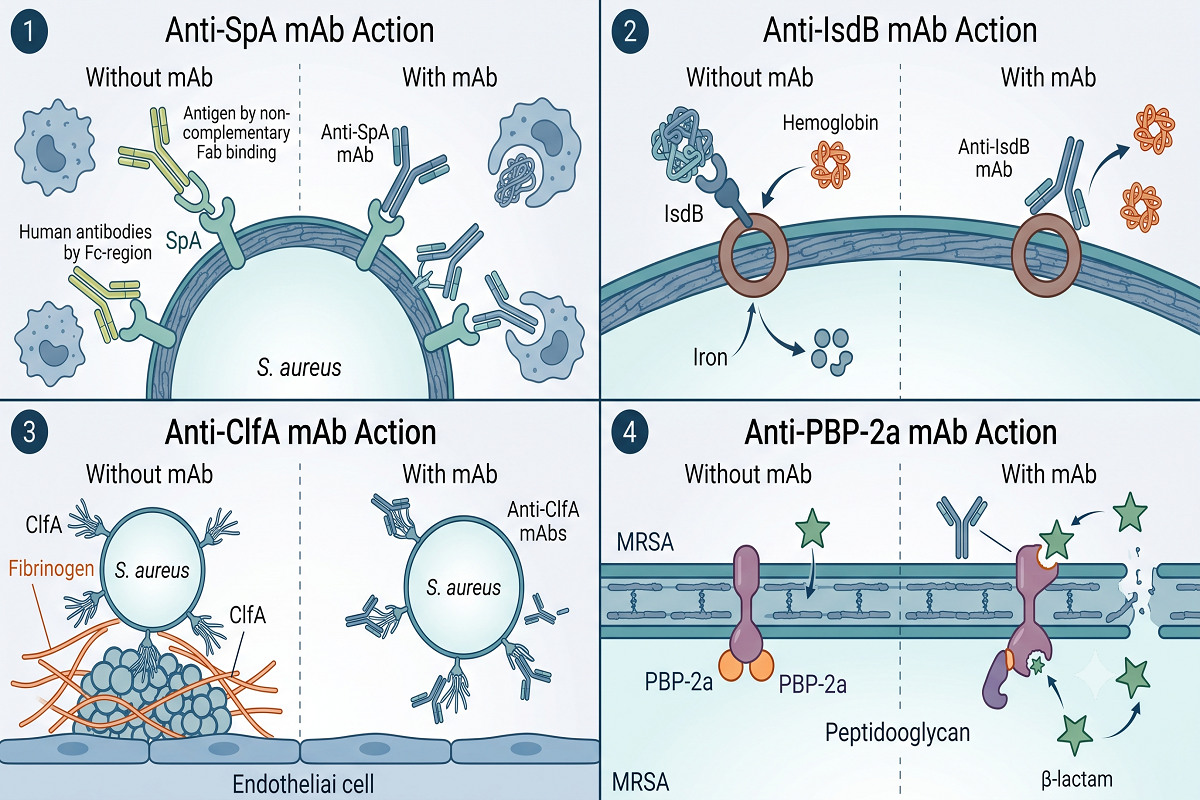
Anti-SpA Antibody Discovery: Neutralizing Immune Evasion
SpA is arguably the most sophisticated immune evasion factor in the S. aureus arsenal. It binds the Fc region of IgG antibodies—preventing opsonophagocytic recognition by neutrophils—and simultaneously cross-links B-cell receptor Fab regions, triggering supraclonal B-cell expansion followed by apoptotic deletion. Through these dual mechanisms, SpA effectively disarms both the humoral and cellular arms of adaptive immunity.
Anti-SpA Antibody Discovery programs face the intrinsic challenge that wild-type SpA is itself immunosuppressive, making conventional immunization approaches ineffective. The solution has been to engineer nontoxigenic SpA variants—such as SpAₖₑₐₐ, in which key glutamine and aspartate residues are mutated to alanine—that lack Fc and Fab binding activity yet retain the structural epitopes needed to elicit cross-reactive antibodies against wild-type SpA. Preclinical studies in murine models have demonstrated that mAbs raised against SpAₖₑₐₐ confer protection against S. aureus challenge by restoring opsonophagocytic clearance. A human anti-SpA mAb (514G3) has advanced to Phase I/II clinical evaluation for S. aureus bacteremia, representing one of the most clinically advanced anti-staphylococcal antibody programs to date.
Anti-IsdB Antibody Discovery: Blocking Iron Acquisition
Iron is an essential nutrient for bacterial growth, and S. aureus has evolved the iron-responsive surface determinant (Isd) system to extract heme-iron from host hemoglobin during infection. IsdB, a ~72 kDa CWA protein, functions as the primary hemoglobin receptor in this pathway. Displaying NEAT (near iron-transport) domains on the bacterial surface, IsdB captures hemoglobin, strips the heme moiety, and relays it through a cascade of Isd proteins (IsdA → IsdC → IsdE/IsdF) for internalization and iron release.
Anti-IsdB Antibody Discovery aims to starve S. aureus of iron at the infection site by blocking the hemoglobin-binding NEAT domains of IsdB. Preclinical studies have shown that anti-IsdB mAbs enhance opsonophagocytic killing in vitro and prolong survival in murine sepsis models. IsdB was also the target of one of the most extensively studied S. aureus vaccine candidates, underscoring its immunologic relevance—although the vaccine’s failure in Phase III trials highlighted the complexity of translating animal model data to human protection. Importantly, IsdB is upregulated under the iron-limited conditions characteristic of the host infection environment, making its expression contextually relevant at the sites where antibody engagement would be most needed.
Anti-ClfA Antibody Discovery: Disrupting Host Adhesion
ClfA is a prototypical MSCRAMM that mediates S. aureus adhesion to host fibrinogen and fibrin, promoting bacterial colonization of damaged endothelial surfaces—a critical early step in the pathogenesis of infective endocarditis and catheter-associated bloodstream infections. Encoded by the highly conserved clfA gene, the 933-amino-acid ClfA protein contains a fibrinogen-binding A region composed of N1, N2, and N3 subdomains, with the N2-N3 cleft serving as the primary ligand-docking site.
Anti-ClfA Antibody Discovery has been pursued through both genetic and immunologic approaches. Passive immunization studies in mice using anti-ClfA antibodies have demonstrated protection against septic arthritis and sepsis-induced mortality, validating ClfA as a bona fide immunotherapeutic target. Notably, tefibazumab (Aurexis), a humanized anti-ClfA IgG1, reached clinical testing for S. aureus bacteremia—making ClfA one of the first S. aureus surface antigens to enter clinical-stage antibody development. Although tefibazumab was ultimately discontinued, it established the feasibility of targeting adhesion factors and generated substantial data on antibody pharmacokinetics and safety in infected patients.
Anti-SasA Antibody Discovery: Targeting a Conserved Surface Protein
SasA is a large (~240 kDa) cell wall-anchored serine-rich repeat protein (SRRP) that has been identified as a virulence determinant in S. aureus intravascular infections. SasA is encoded in virtually all clinical S. aureus isolates examined, and the non-repeat region (NRR)—spanning approximately 500 amino acids—displays over 90% sequence conservation across diverse sequence types.
Anti-SasA Antibody Discovery has been advanced by the characterization of mAb 2H7, a murine IgG1 that binds the conserved NRR domain. In preclinical evaluations, 2H7 achieved 60–70% survival in murine sepsis and peritonitis models against both USA300 (community-associated MRSA) and ST239 (hospital-associated MRSA) strains, with opsonophagocytic killing rates of approximately 60% in vitro and significant reductions in bacterial burden across target organs. These results are notable because they demonstrate cross-strain protection—a critical requirement for any anti-staphylococcal antibody that must contend with the genomic plasticity of S. aureus. The data also suggest that anti-SasA mAbs may work synergistically with anti-SpA and toxin-neutralizing antibodies in combination regimens.
Anti-PBP-2a Antibody Discovery: Countering Antibiotic Resistance
PBP-2a is unique among S. aureus surface antigens in that it is not an endogenous virulence factor but an acquired antibiotic resistance determinant, encoded by the mecA gene located on the staphylococcal cassette chromosome mec (SCCmec). This 78 kDa transpeptidase exhibits markedly reduced affinity for virtually all β-lactam antibiotics, enabling MRSA to continue peptidoglycan cross-linking even when native PBPs are fully inhibited. Structurally, PBP-2a is anchored externally to the bacterial membrane and comprises four domains: a transmembrane anchor, an N-terminal extension, a non-penicillin-binding domain, and a C-terminal transpeptidase domain that catalyzes the DD-transpeptidation reaction essential for cell wall integrity.
Anti-PBP-2a Antibody Discovery represents a fundamentally different strategy from the antigen targets discussed above. Rather than neutralizing a virulence mechanism, anti-PBP-2a antibodies aim to resensitize MRSA to β-lactam antibiotics by blocking the transpeptidase activity that confers resistance. A monoclonal antibody against PBP-2a developed via hybridoma technology (patent WO/2011/017791) has been applied in in vitro diagnostic products for MRSA identification, and the concept of anti-PBP-2a passive immunization combined with β-lactam therapy has been proposed as a potential treatment strategy. By restoring the efficacy of inexpensive, well-characterized β-lactam antibiotics against MRSA, this approach could address both the clinical and health-economic dimensions of the resistance crisis.
Practical Considerations for Multi-Antigen Antibody Discovery
The preclinical data across these five targets collectively illustrate a central lesson from two decades of anti-staphylococcal antibody research: single-antigen approaches may be insufficient against a pathogen with redundant virulence and immune evasion strategies. The failure of several clinical-stage antibody programs—despite compelling preclinical efficacy—has driven a shift in thinking toward multi-antigen combination strategies. An ideal antibody cocktail might pair an anti-SpA component to restore humoral immunity with one or more anti-adhesion mAbs (anti-ClfA, anti-SasA) and, in MRSA settings, an anti-PBP-2a component to reverse β-lactam resistance.
The antibody-antibiotic conjugate (AAC) platform offers an additional dimension by covalently linking anti-staphylococcal mAbs to antibiotic payloads, enabling targeted delivery of bactericidal agents to the bacterial surface or even into the intracellular compartment where S. aureus can persist. For research teams developing antibody-based interventions against S. aureus, access to integrated discovery platforms—spanning antigen design, immunization, hybridoma or display-based screening, lead characterization, and functional validation in relevant in vitro and in vivo models—can substantially accelerate the transition from target identification to lead candidate selection. Tailored preclinical CRO partnerships offer the flexibility to support individual workflow steps or full-program execution, depending on a team’s internal capabilities and project stage.
Conclusion
Antibody discovery against S. aureus surface antigens addresses the pathogen at multiple vulnerable points: immune evasion (SpA), iron acquisition (IsdB), host adhesion (ClfA), surface integrity (SasA), and antibiotic resistance (PBP-2a). Each target offers distinct mechanistic advantages, and the most promising path forward likely involves rational combinations that simultaneously neutralize multiple pathogenic pathways. As antimicrobial resistance continues to erode the efficacy of conventional antibiotics, antibody-based strategies—including multi-target cocktails and AACs—represent a critical component of the next-generation anti-infective pipeline. For research teams advancing S. aureus antibody programs, partnering with an experienced preclinical CRO can provide the specialized expertise and integrated workflows needed to navigate the complexities of anti-bacterial antibody discovery from antigen design through lead optimization.
FAQ
Q: What is Staphylococcal protein A (SpA) and why is it an important antibody target?
A: SpA is a cell wall-anchored S. aureus virulence factor that binds the Fc region of IgG antibodies, preventing opsonophagocytic clearance, and cross-links B-cell receptor Fab regions to induce B-cell apoptosis. Neutralizing SpA with specific antibodies can restore both humoral immunity and phagocytic bacterial clearance, making it a foundational component of multi-antigen anti-staphylococcal antibody strategies.
Q: How does targeting IsdB differ from targeting adhesion factors like ClfA?
A: Anti-IsdB antibodies block the bacterial hemoglobin receptor, depriving S. aureus of the iron required for growth at the infection site. Anti-ClfA antibodies, by contrast, prevent bacterial attachment to host fibrinogen and fibrin on damaged vascular surfaces. These represent complementary rather than redundant mechanisms—one targeting nutrient acquisition, the other targeting tissue colonization.
Q: Why are anti-PBP-2a antibodies considered a different class of anti-S. aureus antibody?
A: Unlike antibodies against SpA, IsdB, ClfA, or SasA—which target virulence factors—anti-PBP-2a antibodies target an antibiotic resistance determinant. By binding PBP-2a on the MRSA surface, these antibodies aim to block the transpeptidase activity that confers β-lactam resistance, potentially restoring the efficacy of β-lactam antibiotics against otherwise resistant strains.
Q: What are the main challenges in developing effective anti-S. aureus antibody therapeutics?
A: The primary challenges include virulence factor redundancy (multiple compensatory pathways), immune evasion factors such as SpA that neutralize antibody effector functions, the genomic diversity of clinical isolates, and the difficulty of translating preclinical efficacy in animal models to human clinical outcomes. These challenges have driven the field toward multi-target combination strategies rather than single-antigen monotherapy.
Q: How can preclinical CRO partnerships support anti-S. aureus antibody discovery programs?
A: Experienced preclinical CROs offer integrated antibody discovery workflows—from computational antigen design and immunization through hybridoma or display-based screening, lead characterization, and functional validation in relevant bacterial challenge models. Flexible engagement models allow research teams to access specialized expertise and infrastructure without building these capabilities internally.