Overview of Circular RNA (CircRNA)
species, stable structure, sequence conservation, and cell- or tissue-specific expression. It is a closed RNA molecule formed by backsplicing, unlike traditional linear RNA. The circRNA does not have a 5'-end cap or a 3'-end poly(A) tail. Through exon circularization or intron circularization, the 3'- and 5'-ends are connected to form a complete circular structure to avoid being degraded by exoribonucleases, making it more stable and conserved than linear RNA. Circular RNA plays a key role in diseases, especially cancer, and has unique advantages in disease diagnosis and the development of therapeutic strategies.
General Knowledge of CircRNAs
Generally speaking, most circRNAs are encoded by known protein genes and consist of one or several exons. circRNAs are mainly produced by head-to-tail back-splicing, but some basic features of linear RNA alternative splicing are also found on circular RNAs, and some circRNAs also contain some exon sequences on RNA. CircRNAs are mainly localized in the cytoplasm because they do not contain cap structures or poly(A) tails. However, some circular RNAs from different sources, such as exon-intron circular RNA (EIciRNA) and circular intronic RNA (ciRNA), will be localized in the nucleus. Possibly due to their unique circular structure, circRNAs are more stable than linear RNAs, which is also the degradation mechanism that makes them resistant to exonucleases and ultimately enables the accumulation of circRNAs in certain cells. In general, circRNAs are expressed at low levels, and interestingly, many circRNAs are upregulated during neurogenesis, and some circRNAs are enriched at synapses. In contrast, circRNAs are downregulated in cancer and some other diseases. It is possible that circRNAs have not reached a plateau due to the faster rate of cell proliferation in these diseases. CircRNAs have so far been shown to be associated with a variety of human diseases, including diabetes, neurological disorders, cardiovascular diseases, chronic inflammation, and cancer. In addition, circRNAs have been found to accumulate during aging.
The Biogenesis of CircRNAs
CircRNAs are covalently closed, endogenous biomolecules in eukaryotes with tissue- and cell-specific expression patterns, whose biogenesis is regulated by specific cis-acting elements and trans-acting factors. CircRNAs are derived from canonical splice sites, and mutational analyses in circRNA expression vectors as well as treatment of HeLa cells with the splicing inhibitor isoginkgetin, which blocks spliceosome assembly, showed that circRNA biogenesis is dependent on the canonical splicing machinery. By contrast, work in Drosophila melanogaster cells has shown that inhibiting the spliceosome by depleting components of the U2 snRNP markedly increases the ratio of circular to linear RNAs. Based on these data, when pre-mRNA processing events are slowed down, nascent RNA can be directed to alternative pathways that facilitate back-splicing. The main hypothesis of back-splicing is that looping of the intron sequences flanking the downstream splice-donor site and the upstream splice-acceptor site brings these sites into proximity. This looping can be mediated by base pairing between inverted repeat elements (such as Alu elements), or by the dimerization of RNA-binding proteins (RBPs) that bind to specific motifs in the flanking introns. However, the biogenesis of many circRNAs is influenced by a combination of cis-acting elements and trans-acting splicing factors, including heterogeneous nuclear ribonucleoproteins (hnRNPs) and SR proteins.
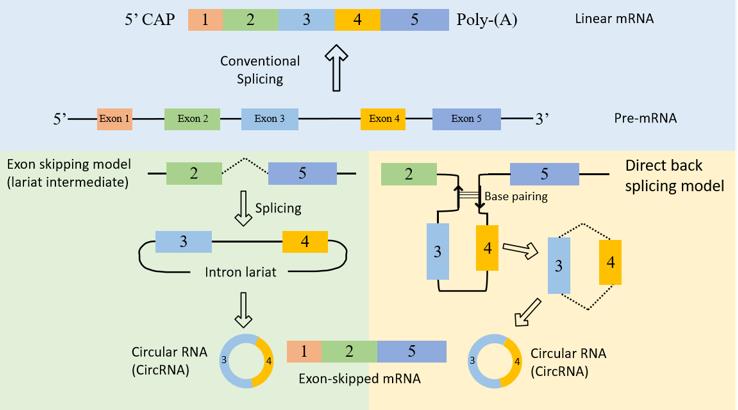 Fig.1 The biogenesis of circRNAs.1
Fig.1 The biogenesis of circRNAs.1
Interestingly, double-stranded RNA (dsRNA)-specific adenosine deaminase (ADAR) enzymes, which prevent the activation of the innate immune system by editing adenosine to inosine in endogenous dsRNA, and ATP-dependent RNA helicase A (also known as DHX9) suppress the biogenesis of circRNAs that rely on base pairing between inverted repeats. Specifically, adenosine-to-inosine editing and the unwinding of dsRNA helical structures prevent the looping of intron sequences. Additionally, lariat formation during exon skipping, an event during which alternative exons are spliced out of the final mRNA product and end up contained within the excised lariat, can lead to the formation of circRNA when the lariat undergoes internal back-splicing. Finally, intronic lariats that escape debranching can lead to the formation of siRNAs.
Cellular Roles of CircRNAs
CircRNAs play an important role in the process of gene expression, from regulating transcription in the nucleus to translation in the cytoplasm. First of all, circRNAs can serve as protein sponges or scaffolds. CircRNA (mainly ciRNA) molecules, located in the nucleus, affect the transcriptional regulation of mother genes and other genes by binding to promoter sites. The second structural analysis found that many circRNAs tend to fold into one to four intramolecular duplexes of 16–26 bp, while less frequently occurring in linear transcripts. This duplex enables circRNAs to bind and inhibit dsRNA-activated protein kinase R (PKR) in cells. Given the central role of PKR in innate immune responses, ectopic expression of circRNAs has been shown to suppress cellular immune responses upon viral infection. Under pathophysiological conditions, the expression levels of circRNAs are reduced in patients with autoimmune diseases. Thirdly, circRNAs can regulate signaling pathways by forming circRNP complexes. Fourth, circRNAs containing multiple miRNA-binding sites can act as competing endogenous RNAs (ceRNAs), thereby modulating the effects of miRNAs on target mRNAs. Fifth, in addition to ceRNAs acting as sponge miRNAs, circRNAs can also directly bind mRNAs to affect gene expression output. Sixth, circRNAs can beat out linear mRNAs by binding to proteins and changing the way mRNAs are translated. This is called competitive binding. At the same time, circRNAs can bind to proteins and act as reaction scaffolds to regulate subsequent biological responses and functions. Finally, in addition to their non-coding role in regulating gene expression, circRNAs have novel roles in coding proteins. They induce the translation of circRNA sequences through the internal ribosome entry site (IRES) sequence or m6A modification in a cap-independent manner.
Preparation of CircRNAs
The efficient synthesis of circRNAs is fundamental to understanding circRNAs and their biomedical applications. Many chemical and intracellular methods have been developed to synthesize circRNAs. Typically, circRNAs are synthesized by synthesizing one or more precursor linear RNAs, followed by chemical or enzymatic ligation-mediated RNA circularization. At present, there are three commonly used methods for the artificial preparation of circular RNAs, including type I intron (T4 bacteriophage or Anabaena) self-cleavage, type II intron self-cleavage, and T4 RNA ligase method. Through sequence optimization design, T4 bacteriophage can be used to prepare circRNA without residues of foreign sequences in vitro. However, the efficiency of preparing circRNA by this method is low, and it cannot meet the needs of industrialization for the time being. CircRNAs can be artificially produced in vitro using autocatalytic splicing reactions from yeast type II introns. The circRNA prepared by this method has no residue of foreign sequences, but the preparation efficiency is low, and it is difficult to carry out industrial preparation at present. The traditional method uses T4 DNA or T4 RNA ligase to realize in vitro circular RNA preparation with the aid of the splint sequence, but the preparation efficiency is low and the purification process is difficult, so it is difficult to realize industrial preparation. More recently, the circularization of RNA without additional splint strands or auxiliary strands has been achieved by computer simulation of the RNA structure and finding the breaks for optimal RNA circularization. This method can significantly inhibit the generation of macromolecular by-products in the circularization process, and allow RNA sequences that are difficult or impossible to circularize to be efficiently circularized. The concept of redesigning the circularization interface sequence is not only suitable for the preparation of T4 ligase but also compatible with the intron self-cleavage system for the efficient preparation of circular RNA in vitro.
Application of CircRNAs
According to current knowledge about circRNAs, there are several remarkable characteristics of circRNAs that make them potential biomarkers for human diseases. The average half-life of circRNAs in plasma exceeds 48 h, much longer than the 10 h of the average value of mRNAs. CicRNAs are suggested to be the most universal molecules distributed in human cells, and in some circumstances, circRNAs are way more abundant than their linear isoforms. CircRNAs are suggested to express in a tissue- and developmental-stage-specific manner, which makes them potential biomarkers of specific diseases. In particular, lots of studies have demonstrated that circRNAs are distinctively expressed in cancerous and non-cancerous tissues. circRNAs are found to be evolutionally conserved in different species, which means some circRNA biomarkers identified in murine models hold the potential to be translated into clinical applications for human beings. Taken together, circRNAs are favorable potential biomarkers for human diseases because of their stability, specificity, and abundance in body fluids.
Reference
- Verduci, Lorena & Strano, Sabrina & Yarden, Yosef & Blandino, Giovanni. (2019). The circ RNA ‐micro RNA code: emerging implications for cancer diagnosis and treatment. Molecular Oncology. 13. 10.1002/1878-0261.12468. Distributed under Open Access license CC BY 4.0, without modification.